
- Title
-
Estrogen defines the dorsal-ventral limit of VEGF regulation to specify the location of the hemogenic endothelial niche
- Authors
- Carroll, K.J., Esain, V., Garnaas, M.K., Cortes, M., Dovey, M.C., Nissim, S., Frechette, G.M., Liu, S.Y., Kwan, W., Cutting, C.C., Harris, J.M., Gorelick, D.A., Halpern, M.E., Lawson, N.D., Goessling, W., North, T.E.
- Source
- Full text @ Dev. Cell
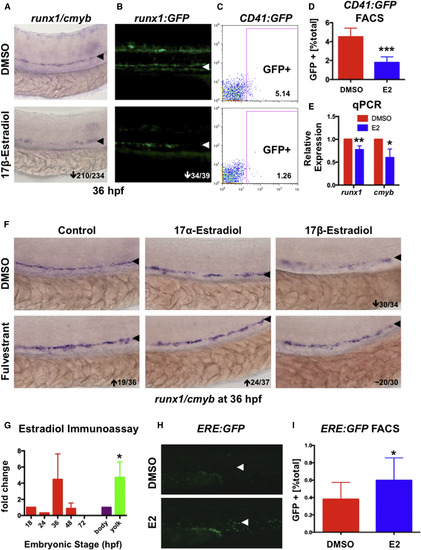
Exposure to E2 Impairs Formation of HSPCs in the AGM (A) E2 exposure from five somites to 36 hpf decreased runx1/cmyb expression in the AGM (210/234). (B) E2 diminished runx1:GFP+ cells in the AGM (34/39). (C) FACS analysis of CD41:GFP embyros (representative samples shown) confirmed that E2 reduced HSCs. (D) CD41:GFP was significantly decreased by E2 (n = 20, two-tailed t test, p < 0.001; error bars indicate SD). (E) qPCR quantified reduced runx1 and cmyb expression by E2 (mean of triplicate experiments ± SEM; one-tailed t test runx∗∗p < 0.01, cmyb∗p < 0.05). (F) The pan-Esr antagonist FULV blocked the effect of E2; the isomer 17α-estradiol (10 μM) had no effect (n ≥ 30/treatment). (G) Estradiol EIA revealed that endogenous estrogens are present during hematopoiesis; E2 was enriched 4.7-fold in the yolk versus body at 18 hpf (mean of triplicate experiments ± SEM; two-tailed t test, ∗p < 0.05). (H) ERE:GFP fish show estrogenic activity in the AGM. (I) GFP expression (as seen in H) was increased 1.57-fold upon E2 exposure (n = 15, two-tailed t test, p < 0.05; error bars indicate SD). The number of embryos with altered expression over the total number analyzed is shown (bottom right). |
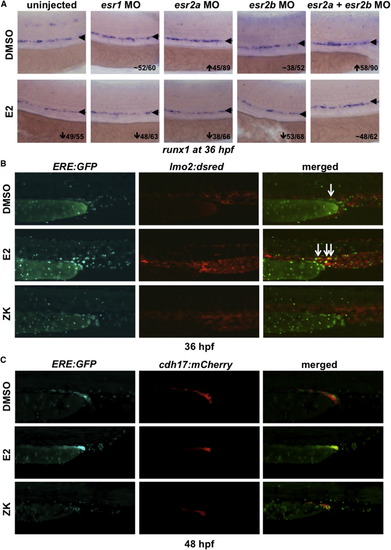
ERE:GFP Embryos Display Estrogen Activity in the Developing AGM (A) MO knockdown showed that the effect of E2 on HSPCs is mediated through esr2a+b; esr1-MO had no impact on runx1/cmyb (n ≥ 30). (B) ERE:GFP embryos show low expression in the trunk vasculature, labeled by lmo2:dsRed; ERE activity expands following E2 exposure and is alleviated by treatment with ZK164015 (n ≥ 20). (C) ERE:GFP embryos exhibit activity in the developing mesonephros, as indicated by cdh17:mCherry at 48 hpf, which responds to altered E2 signaling (n ≥ 20). |
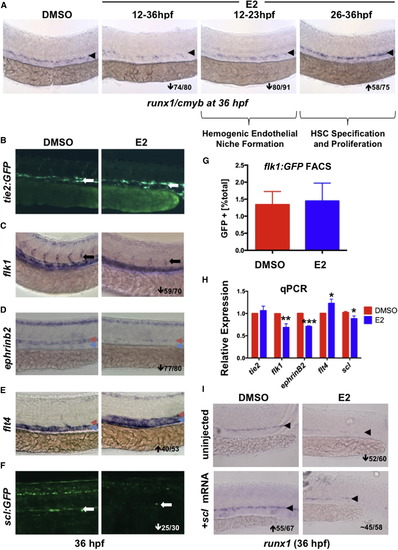
E2 Exposure Impairs Formation of the Hemogenic Endothelial Niche (A) E2 had a biphasic effect on HSPCs: treatment during hemogenic niche specification (12–23 hpf) decreased runx1, whereas exposure during HSC specification and budding (26–36 hpf) increased HSPCs (n ≥ 75/treatment). (B) Treatment with E2 had no impact on axial vessel formation as assessed by tie2:GFP (n ≥ 20). (C) E2 disrupts ISV formation as indicated by flk1 WISH (59/70). (D) Arterial ephrinb2 was robustly decreased by E2 (77/80); red arrowheads indicate artery, and blue arrowheads indicate vein. (E) Venous flt4 was increased after E2 exposure (40/53). (F) scl:GFP was reduced by E2 treatment (25/30). (G) FACS of flk1:GFP+ embryos (n = 32) revealed no change in endothelial cell number by E2 (error bars indicate SD). (H) qPCR confirmed decreased expression of flk1, ephrinB2, and scl following E2 treatment (mean of triplicate experiments ± SEM; one-tailed t test; flt4∗p < 0.05, flk1∗∗p < 0.01, and ephrinB2∗∗∗p < 0.001). (I) Injection of scl mRNA increased runx1 expression in the AGM and rescued loss following E2 treatment (n > 55 per treatment). |
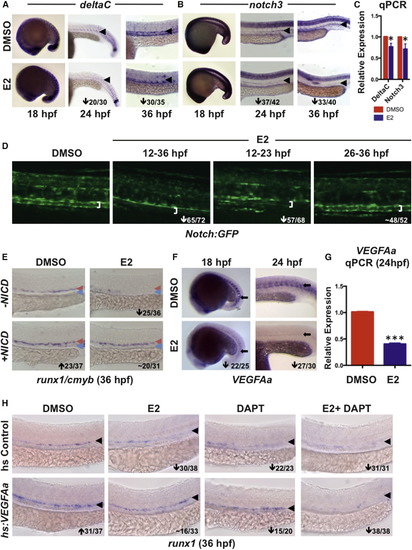
Exogenous E2 Signaling Disrupts VEGF/NOTCH Cascades in the AGM (A) E2 exposure had no impact on deltaC expression by WISH at 18 hpf but strongly reduced arterial expression at 24 and 36 hpf (n > 30/treatment); an arrowhead indicates artery. (B) Expression of notch3 was similarly regulated by E2 at 18, 24, and 36 hpf (n > 30/treatment); an arrowhead indicates artery. (C) qPCR confirmed E2-mediated reductions in deltaC and notch3 expression at 36 hpf (mean of triplicate experiments ± SEM; one-tailed t test deltaC, notch3∗p < 0.05). (D) Decreased Notch:GFP was observed in the artery (white bracket indicates artery walls) after E2 treatment from 12–36 or 12–23 hpf; no change occurred with exposure from 26–36 hpf (n ≥ 52). (E) NICD induction increased expression of runx1/cmyb and rescued HSPCs in E2-treated embryos (n ≥ 31/treatment). (F) Expression of VEGFAa was strongly decreased in zebrafish following E2 treatment at both 18 and 24 hpf (n ≥ 25/treatment); an arrow points to somite staining. (G) qPCR confirmed that expression of VEGFAa was decreased at 24 hpf (mean of triplicate experiments ± SEM; one-tailed t test ∗∗∗p < 0.001). (H) Induction of VEGFAa increased runx1 expression and rescued the effect of E2 exposure; treatment with DAPT decreased expression of runx1 and blocked the VEGF-mediated rescue of HSPCs (n ≥ 20/treatment); an arrowhead indicates artery. EXPRESSION / LABELING:
PHENOTYPE:
|
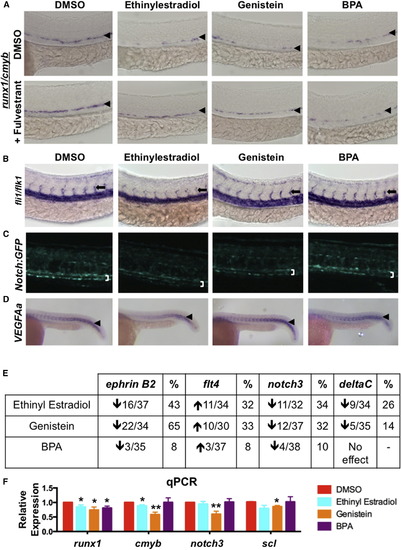
Xenoestogens Partially Replicate the Effects of E2 on HSPC Formation (A) Exposure to xenoestrogens ethinyl estradiol (EE), genistein (GEN), or Bisphenol A (BPA) decreased runx1/cmyb expression and could be partially blocked by cotreatment with FULV (n ≥ 25/treatment). (B) Treatment with GEN reduced the expression of fli1/flk1 in the intersomitic vessels (n ≥ 25/treatment); an arrow points to ISVs. (C) Treatment with GEN (15/25) and EE (12/23) decreased Notch:GFP expression. (D) VEGFAa was noticeably decreased by EE (15/26); arrowheads indicate somite region. (E) Summary of changes in the expression of markers of vascular identity ephrinB2, flt4, notch3, and deltaC following treatment with XEs (n ≥ 25). (F) qPCR confirmed decreases in runx1, cmyb notch3, and scl by XE exposure (mean of triplicate experiments ± SEM; one-tailed t test ∗p < 0.05; ∗∗p < 0.01). EXPRESSION / LABELING:
PHENOTYPE:
|
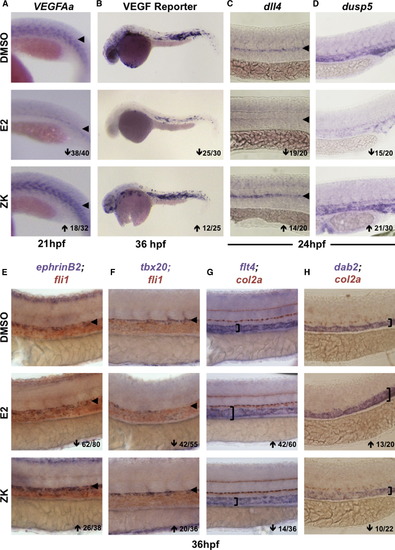
Blocking Esr Signaling Increases VEGF Activity and Alters Trunk Vasculature Patterning (A) VEGFAa expression was enhanced after inhibition of endogenous estrogen activity via the pan-Esr antagonist ZK164015 (18/32). (B) gfp expression in VEGF:GFP reporter embryos was reduced by E2 and enhanced by ZK (n > 25). (C) The VEGF target gene dll4 was decreased after E2 exposure but increased by ZK (n > 20). (D) dusp5 levels were reduced by E2 treatment but elevated after Esr antagonism (n > 20). (E) Arterial ephrinB2 expression in the fli1+ vasculature was enhanced after ZK-mediated inhibition of estrogen signaling (26/38). (F) tbx20 expression in the roof of the aorta was substantially increased by ZK (20/36). (G) Venous flt4 was expanded dorsally toward the col2a+ hypocord by E2 but reduced in expression by ZK exposure (14/36). (H) The venous marker dab2 was similarly regulated as determined by WISH (n > 20). EXPRESSION / LABELING:
PHENOTYPE:
|
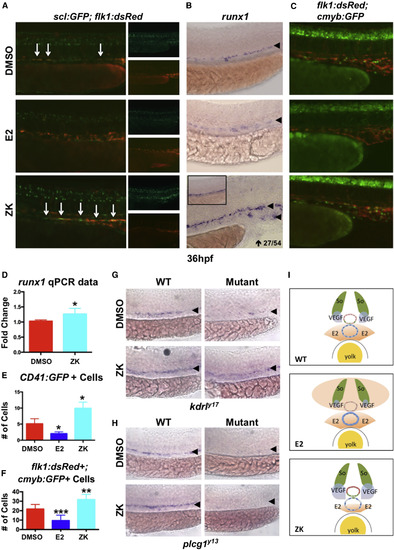
E2-Mediated VEGF Antagonism Regulates Hemogenic Endothelial Specification (A) Hemogenic endothelium, as assessed by the dual expression of scl:GFP and flk1:dsRed, was reduced by exogenous E2 treatment but was enhanced following Esr antagonism by ZK (n > 15). (B) runx1 expression increased (27/54) after estrogen receptor inhibition, with mislocalization to the vein (10/54). (C) Production of flk1:dsRed+; cmyb:GFP+ HSCs was enhanced by ZK (n ≥ 10). (D) Quantification of changes in runx1 expression (mean of triplicate experiments ± SEM; one-tailed t test ∗p < 0.05). (E) Quantification of the number of CD41+ cells after E2 treatment and antagonism (one-tailed t test ∗p < 0.05; error bars indicate SD). (F) The number of flk1:dsRed+; cmyb:GFP+ HSCs was decreased by E2 and enhanced after ZK treatment (one-tailed t test ∗∗p < 0.01, ∗∗∗p < 0.001; error bars indicate SD). (G) kdrly17 mutant embryos reveal loss of runx1, which can be partially restored by reduction of E2-mediated VEGF antagonism after ZK treatment (n > 20/condition). (H) WISH analysis in plcg1y13 mutant embryo shows complete loss of runx1 expression in the presence or absence of ZK (n > 20/condition). (I) Model: in wild-type embryos (upper panel), opposing gradients of E2 and VEGF specify the arterial/venous boundaries of hemogenic endothelium and subsequent emergence of HSPCs (red circle, artery; blue circle, vein). Excess E2 (middle) disrupts this gradient, further antagonizing VEGF and causing artery, hemogenic endothelium, and HSPCs to fail to form properly. In contrast, inhibition of esr activity (lower panel) allows the range of VEGF regulation to increase, resulting in ectopic HE specification and HSPC production in the vein. |
Reprinted from Developmental Cell, 29, Carroll, K.J., Esain, V., Garnaas, M.K., Cortes, M., Dovey, M.C., Nissim, S., Frechette, G.M., Liu, S.Y., Kwan, W., Cutting, C.C., Harris, J.M., Gorelick, D.A., Halpern, M.E., Lawson, N.D., Goessling, W., North, T.E., Estrogen defines the dorsal-ventral limit of VEGF regulation to specify the location of the hemogenic endothelial niche, 437-53, Copyright (2014) with permission from Elsevier. Full text @ Dev. Cell