
- Title
-
The Evolution of Feeding Mechanics in the Danioninae, or Why Giant Danios Don't Suck Like Zebrafish
- Authors
- Conith, M.R., Ringo, D., Conith, A.J., Deleon, A., Wagner, M., McMenamin, S., Cason, C., Cooper, W.J.
- Source
- Full text @ Integr Org Biol
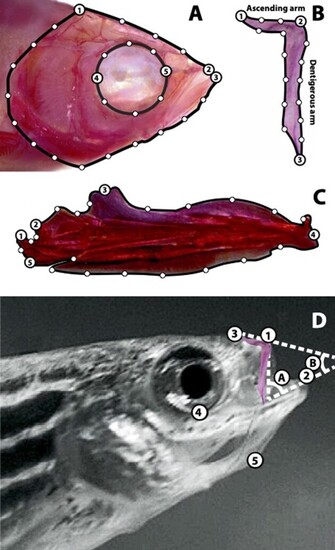
Shape and motion data collection. A. Homologous anatomical landmarks (LM) used in skull shape analyses: (1) most posterio–dorsal point on the parietal bone; (2) anterior tip of the premaxilla; (3) anterior tip of the dentary; (4) posterior point on the eye socket in line with the parasphenoid; and (5) anterior point on the eye socket in line with the parasphenoid. Semilandmarks were evenly spaced along the outline of the skull between LM 1 and 2, 1 and 3, 4 and 5, and 5 and 4. B. Homologous anatomical LM used in premaxilla shape analyses: (1) tip of the ascending arm; (2) anterior tip of the premaxilla (see panel D for the location of the premaxilla within the skull); (3) and tip of the dentigerous arm. Semilandmarks were evenly spaced along the outline of the premaxilla between LM 1 and 2, 2 and 3, and 1 and 3. C. Homologous anatomical LM used in mandible shape analyses: (1) posterio–dorsal extent of the lower jaw joint (quadrate–articular joint); (2) anterio–dorsal extent of the lower jaw joint; (3) tip of the primordial process of the articular; (4) anterior tip of the dentary; and (5) posterior tip of the angular. Semilandmarks were evenly spaced along the outline of the mandible between LM 1 and 2, 2 and 3, 3 and 4, 4 and 5, and 1 and 5. D. LM and angles used in motion analyses: (1) anterior tip of the premaxilla; (2) anterior tip of the dentary; (3) a stationary point on the dorsal surface of the anterior head; (4) ventral-most point of the eye socked; (5) anterior tip of the hyoid series (basihyal); (A) gape angle; and (B) angle of jaw-protrusion direction. |
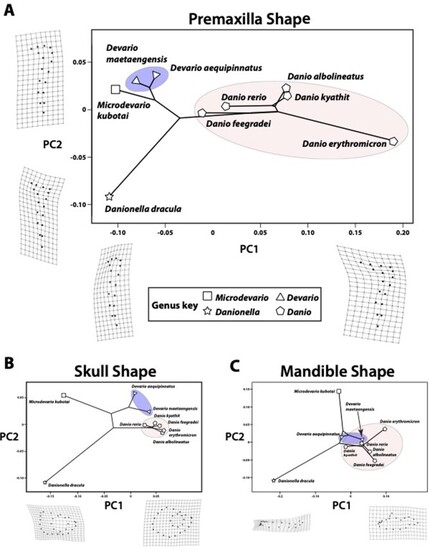
PCA-based phylomorphospaces for premaxilla, skull, and mandible shape diversity in the Danioninae. A. Premaxilla shape diversity. Deformation grids depict the shapes associated with high and low PC scores on both axes. B. Skull shape diversity. Deformation grids depict the shapes associated with high and low PC1 scores. C. Mandibular shape diversity. Deformation grids depict the shapes associated with high and low PC1 scores. |
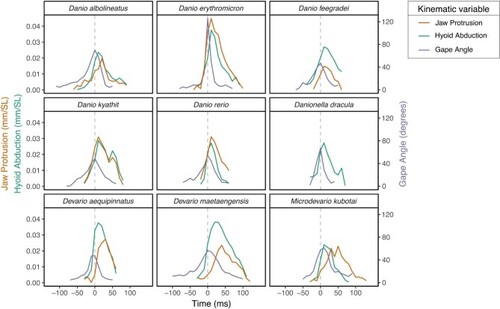
Time series for jaw protrusion, hyoid depression, and gape angle during feeding strikes for nine danionine species. Species means are shown. Horizontal bars indicate the time elapsed between maximum hyoid depression and maximum jaw protrusion. There are no horizontal bars for Danio erythromicron, D. feegradei, D. quagga, or D. rerio because these maxima occurred simultaneously. |
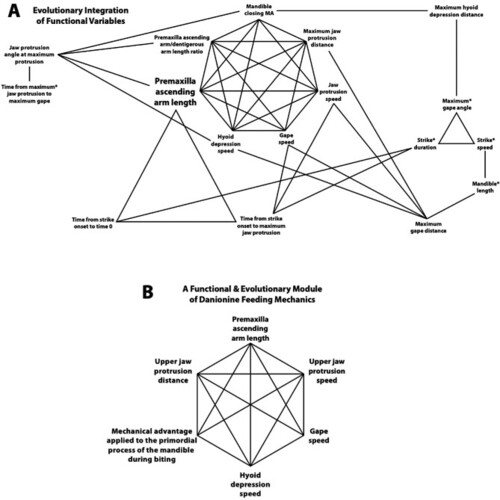
Evolutionary patterns of integration among variables that describe the functional morphology of feeding in the Danioninae. A. Connections between variables indicate that phylogenetic generalized least squares (PGLS) analyses showed their evolution to have been significantly correlated (see |

The evolution and development of danionine feeding mechanics. A. Tracings of two danionine species taken from high-speed videos at ∼80–85% maximum jaw protrusion. They typify two feeding strategies that employ alternative jaw-protrusion techniques and different premaxilla shapes: Devario aequipinnatus (typical of Devario and Microdevario species); Danio erythromicron (typical of Danio species). B. Chronogram showing the relationships and divergence times of the 9 species examined here. Body sizes are relative. The emergence of three feeding strategies associated with differences in jaw protrusion are marked. C. Variation in premaxilla shape among the Danioninae. There is limited variation in ascending arm length (all very short) in the DM clade. Ascending arms are longer in the Danio clade and there is greater length variation. D. Premaxilla development in the Danioninae. The ascending arm elongates during post-metamorphic development in wild-type zebrafish when thyroid hormone (TH) is present. Ascending arms do not elongate in hypothyroid zebrafish (-TH; Galindo et al. 2019). Post-metamorphic ascending arm elongation does not occur in Devario aequipinnatus. E. Mandible elongation in zebrafish mutants with congenitally elevated (+TH) levels. F. Changes in the ratio of premaxilla ascending arm length to dentigerous arm length evolved significantly faster among fishes that use jaw protrusion for suction production (Danio) than among those that use it for prey retention (DM). Changes in the relative timing of maximum jaw protrusion and maximum gape evolved significantly faster among fishes that use jaw protrusion for suction production (Danio) than among those that use it for prey retention (DM). |