
- Title
-
Leveraging Zebrafish to Study Retinal Degenerations
- Authors
- Angueyra, J.M., Kindt, K.S.
- Source
- Full text @ Front Cell Dev Biol

Structure of the zebrafish eye and retina. |
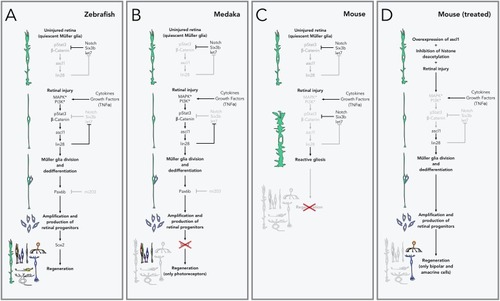
Pathways to retinal regeneration. |

Tools to study retinal circuits. |