
- Title
-
Development of a straight vertebrate body axis
- Authors
- Bagnat, M., Gray, R.S.
- Source
- Full text @ Development
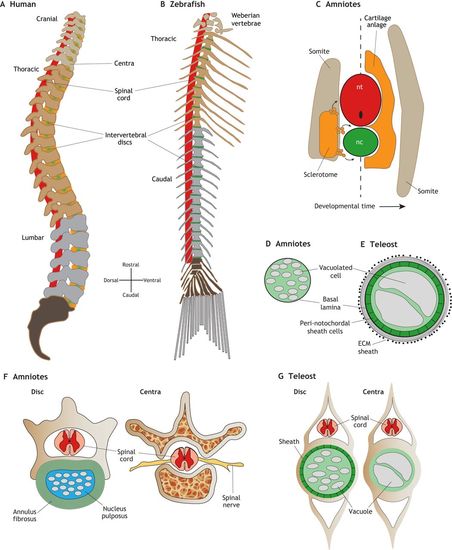
Development and morphology of the spine in amniotes and zebrafish. (A) Lateral view schematic of a human adult spine. The intervertebral discs (green/orange) are largely derived from the cartilaginous anlage and notochord in mammals. (B) Lateral view schematic of a zebrafish adult spine. The intervertebral discs (green) are largely derived from the notochord in zebrafish. (C) Transverse view schematic of re-segmentation of the somite-derived sclerotome (orange) tissue which migrates out from the somite (left side of dashed line) to form the cartilage anlage, which wraps the spinal cord and notochord (right side of dashed line). The myotome region of the somite give rise to the axial muscles (tan). (D) Schematic of an amniote notochord in cross section, showing vacuolated cells and a basal lamina sheath. (E) Schematic of a teleost notochord in cross section, showing a larger notochord with large vacuolated cells enclosed by a peri-notochordal sheath cell epithelium, and a lamellar ECM sheath. (F) Cross-sections of mouse spine highlighting the multi-lamellar annulus fibrosus fibrocartilage layer (green), surrounding the nucleus pulposus (light blue), which contains notochord-derived cell lineages, the spinal cord (pink/red) and spinal nerves (yellow), and the trabecular bony regions. (G) Cross-sections of zebrafish spine showing the intervertebral disc (Disc) region, highlighting the sheath and small fragmented vacuolated cells that form the zebrafish disc and the large vacuolated cells within the vertebral body region. |
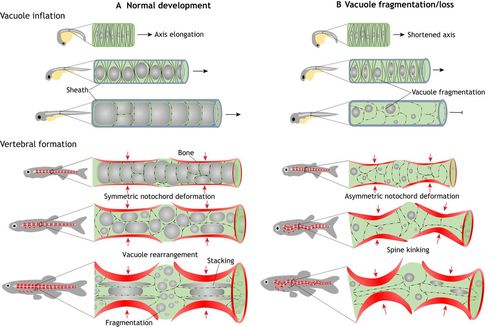
Role of notochord vacuolated cells in axis elongation and spine formation. (A) During normal development, as vacuolated cells nucleate and inflate their large fluid filled vacuole (gray spheres) in an anterior-to-posterior direction within the semi-rigid sheath they generate hydrostatic pressure that pushes axis elongation. Later in development, vertebral bone (red) is deposited around the notochord in a segmented fashion, creating an alternating pattern of vertebral and intervertebral domains. As the vertebrae grow concentrically, the large vacuoles are able to absorb the compressive force, allowing symmetrical bone growth. During this period, some vacuolated cells are stacked under the growing bone, whereas in other cells, vacuoles fragment and the cells localize to the IVD domains. This process allows for symmetric bone growth, IVD formation and a straight spine formation. (B) When vacuoles fragment prematurely or are lost, internal notochord pressure is lost, leading to shortening of the embryonic axis. Vacuole loss also causes the notochord to lose its ability to absorb the force of the growing vertebrae evenly and locally. As vertebral bone grows the notochord is deformed asymmetrically, causing vertebral malformation and kinking of the spine axis. |
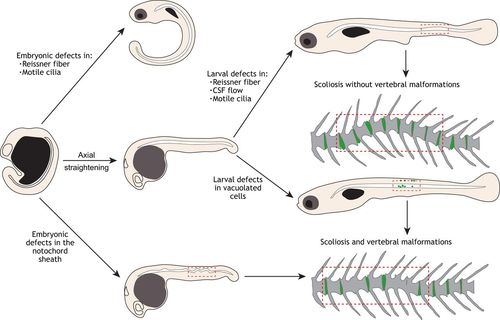
Processes regulating axial straightening and notochord integrity underlie defects of spine morphogenesis. Beginning at the curved tailbud stage, generating a straight body axis requires the function of motile cilia components, which drives the formation of the Reissner fiber (top). Once axial straightening occurs, defects of the Reissner fiber, disrupted motile cilia or CSF flow, which are revealed during larval development in zebrafish, can lead to more subtle curvatures of the notochord. Alternatively, defects in the notochord sheath or in vacuolated cell morphogenesis can contribute to severe bends or kinks of the notochord, which directly arise into regions of vertebral malformations and fusion, and sometimes scoliosis. |
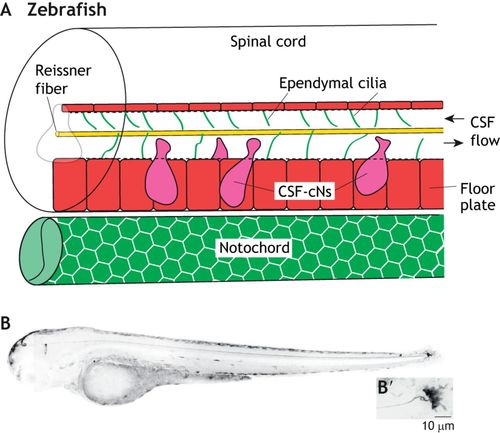
The integration of the Reissner fiber and CSF-resident components. (A) Schematic of the larval zebrafish spinal cord and notochord, showing the close associations of the Reissner fiber (yellow) with ependymal motile cilia (green lines) and CSF-cNs (magenta). The floor plate is shown in red. (B) Inverted grayscale confocal image of a 3 days post-fertilization larval scospondin-GFPut24 knock-in line in a lateral view. (B’) High magnification image of the terminal ampulla region. Taken from Troutwine et al., 2019 preprint. |