
- Title
-
Problems in Fish-to-Tetrapod Transition: Genetic Expeditions Into Old Specimens
- Authors
- Wood, T.W.P., Nakamura, T.
- Source
- Full text @ Front Cell Dev Biol

The evolution of cranial dermal bones and the developmental mechanisms. |
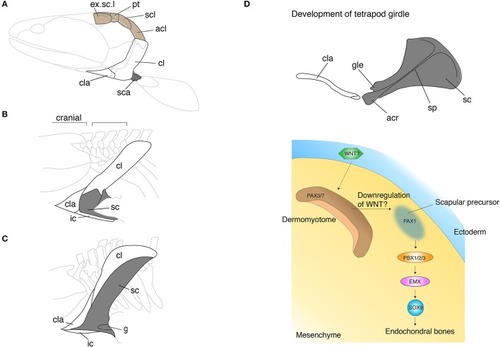
The skeletal shift from dermal to endochondral bones in the pectoral shoulder girdles. |

The fin-to-limb transition and their developmental basis. |