
Fig. S7
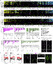
Comparison of Brain Activity and Behavioral Datasets, Related to Figure 2 and Figure 4 (A) Comparison of baseline behavioral data across multiple time windows (Figure 2) with brain activity changes in the four major regions. Stimulus-driven behaviors (prepulse, dark flash response, etc) are not comparable to the baseline phospho-Erk data. There are several caveats to consider when comparing these two datasets. The behavioral data was collected on larvae in 96-well plates, isolated from interaction with any other larvae and constrained in their movement, while phospho-Erk staining was completed on a group of larvae in a large (150 mm) Petri dish. It is likely that the differences in these conditions would result in different brain states. The phospho-Erk dataset also represents only a single time point, the afternoon of 6 dpf, while the behavioral data was collected across two nights and days. Both the phospho-Erk and behavioral analyses only explore a small and relatively non-overlapping part of the possible behavioral landscape, and a more appropriate assessment would be to compare these two types of data collected under the same conditions for specific behaviors of interest. Further, finding similar differences in brain activity in a region does not necessarily mean that the exact same neurons are affected. (B) To search for connections between brain activity and behavior in an unbiased and quantitative way, we developed a procedure to compare the merged p values from baseline behavioral assays with the region-specific brain activity signal. Two relationships remained convincing after qualitative assessment (open circles) of the results: a relationship between reduced frequency of motion and a reduced activity in the telencephalon (C), and a relationship between increased activity in the rhombencephalon and increased preference for the well edge (D). Briefly, we binarized both the brain activity and behavioral data and conducted Fisher exact enrichment analyses, followed by combining the resulting p values for all time windows to look for relationships that were consistent across time and therefore more likely real. For the brain activity data, we focused on the 61 largest regions based on their size in pixels, and normalized the total number of pixels with signal by dividing by the region size. Regions with less than 1% activity were designated as 0 and regions with greater than 1% signal were designated as 1. This binarization was not optimal and certainly resulted in false positive designations of signal (filled-in circles represent contributions to quantitative significance that were potentially false designations, compared to open circles where binarization more accurately represented the data). For the behavioral data, any assay with a significant merged p value (< 0.05) was designated as 1. Following initial enrichment analyses across a matrix of the 61 regions and 45 baseline time windows, the resulting Fisher p values were combined across the time windows using Fisher’s method. The most significant combined p values are shown for the comparisons of both increased behavior and decreased with increased and decreased binarized brain activity signal. If a comparison is not shown, no combined p values of substantial significance were identified. (C) Brain activity maps for the forebrains of eight mutants that have both convincing telencephalon activity reductions and a consistent reduction in baseline frequency of motion (open circles, A). (D) Three examples of mutants with an increased preference for the well edge. Correspondingly, we observed substantial increases in brain activity in these mutants, specifically in the hindbrain region for elfn1(see also Figure S5 brain activity map for only elfn1a) and hcn1. Additional examples are highlighted with open circles (A) and are available to view as stacks on stackjoint.com/zbrain. Intriguingly, several of these mutants have also been implicated in epilepsy (references in Table S1), another neuropsychiatric disease that may share genetic underpinnings with schizophrenia. |
Reprinted from Cell, 177(2), Thyme, S.B., Pieper, L.M., Li, E.H., Pandey, S., Wang, Y., Morris, N.S., Sha, C., Choi, J.W., Herrera, K.J., Soucy, E.R., Zimmerman, S., Randlett, O., Greenwood, J., McCarroll, S.A., Schier, A.F., Phenotypic Landscape of Schizophrenia-Associated Genes Defines Candidates and Their Shared Functions, 478-491.e20, Copyright (2019) with permission from Elsevier. Full text @ Cell