
- Title
-
Hedgehog signaling induces arterial endothelial cell formation by repressing venous cell fate
- Authors
- Williams, C., Kim, S.H., Ni, T.T., Mitchell, L., Ro, H., Penn, J.S., Baldwin, S.H., Solnica-Krezel, L., and Zhong, T.P.
- Source
- Full text @ Dev. Biol.
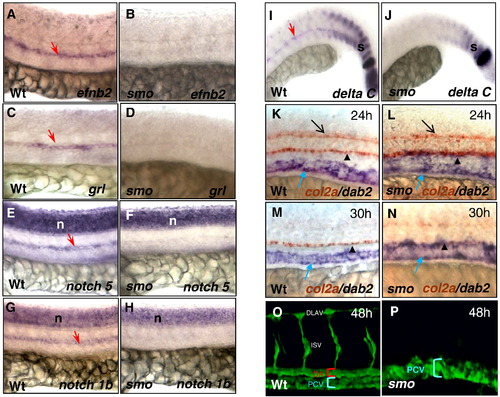
smo mutants display expansion of the posterior cardinal vein and absence of the dorsal aorta. (A–J) Lateral views displaying absent expression of efnb2 (A, B), grl (C, D), notch5 (E, F), notch1b (G, H) and deltaC (I, J) in smo mutants, compared to wild-type embryos. (K–N) Double in situ hybridization exhibiting the expansion of dab2 venous domain and col2a expression in the neural tube and the hypocord in smo mutants (L, N) compared to wild-type embryos (K, M). (O, P) Confocal microscopy depicting the dorsal aorta and the posterior cardinal vein in wild-type embryos [Tg(flk:EGFP)] (O); and absence of the dorsal aorta and expansion of the posterior cardinal veins in smo mutants [smohi640/smohi640; Tg(flk:EGFP)] (P). Red arrow: dorsal aorta. Blue arrow: cardinal vein. Black arrow: floor plate. Black arrowhead: hypocord. s: somite. n: neural tube. DA: dorsal aorta. PCV: posterior cardinal vein. DLAV: dorsal longitudinal anastomotic vessel. |
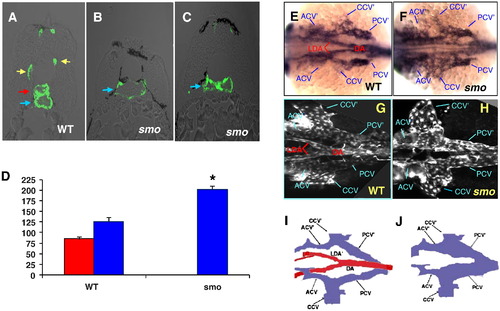
smo mutation causes loss of the aortic bifurcation. (A–C) Transverse section analyses showing normal lumen sizes of the dorsal aorta and the posterior cardinal vein in wild-type embryos (A); single enlarged cardinal vein (B) and collapsed cardinal vein in smo mutants (C). (D) Bar chart depicting perimeter measurements of lumen sizes of the dorsal aorta and the cardinal vein in wild-type embryos and smo mutants. Lumen sizes were measured in 15 transverse sections derived from three wild-type and three smo mutant embryos, respectively. Error bars indicate standard deviation and asterisks indicate statistical significance between wild-type and smo data (P < 0.01). (E–H) In situ hybridization of flk1 at 24 hpf (E, F) and confocal microscopy in Tg(flk1:EGFP) embryos at 48 hpf (G, H) exhibiting loss of the aortic bifurcation and expansion of bilateral cardinal veins in smo mutants (F;H), compared to the normal aortic bifurcation and bilateral cardinal veins in wild-type embryos (E;G). (I, J) Schematics depicting the aortic bifurcation flanked by bilateral cardinal veins in wild-type embryos, compared to enlarged bilateral cardinal veins in smo mutants. Red arrow: aorta. Blue arrow: vein. Yellow arrow: intersomitic vessel. DA: dorsal aorta. LDA: lateral dorsal aorta. acv: anterior cardinal vein. ccv: common cardinal vein. pcv: posterior cardinal vein. LDA′, acv′,ccv′, pcv′: denote the corresponding vessel on the other side of the embryo. |
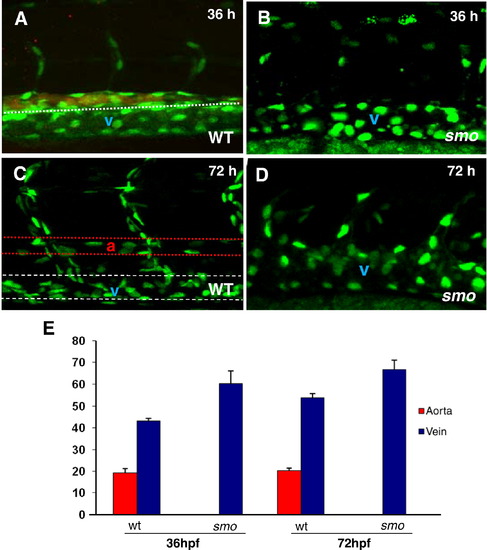
The increase of venous cells is equivalent to loss of arterial cells in smo mutants. (A–D) Confocal optics displaying individual nuclei of endothelial cells in the dorsal aorta (A, B stained with efnb2 antibody, the posterior cardinal vein and intersomitic vessels in wild-type embryos [Tg(fli:EGFP-nuc)] at 36 hpf (A) and 72 hpf (C),as compared to the increased endothelial nuclei in enlarged cardinal veins in smo mutants [smohi1640/smohi1640;Tg(fli:EGFP-nuc)] at 36 hpf (B) and 72 hpf (D). (E) Bar chart depicting number of arterial and venous cells in wild-type embryos compared to smo mutants. At 36 hpf, arterial cell number: 19.2 ± 1.7 (WT), 0 (smo); Venous cell number: 43.2 ± 1.3 (WT), 60.3 ± 2.4 (smo). At 72 hpf, arterial cell number: 20.2 ± 1.3 (WT), 0 (smo); Venous cell number: 54 ± 1.6 (WT), 66.8 ± 3.4 (smo). Endothelial cells were counted from 12 lateral sections derived from 6 wild-type embryos and 6 smo mutants. Each section covers four segment lengths of intersomitic vessels along the dorsal aorta and the posterior cardinal vein in the middle trunk above the yolk extension region. a: aorta. v: vein. |
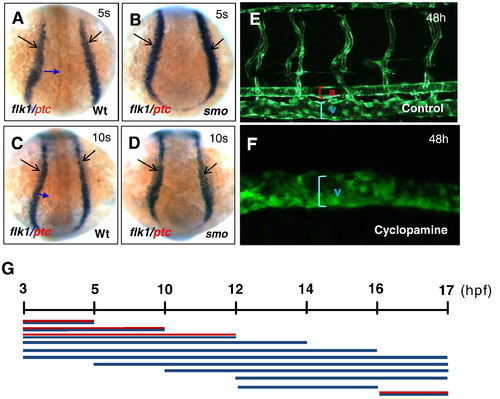
Temporal development stages when Hh signaling affects artery–vein specification. (A–D) Dorsal views displaying comparable levels of flk1-positive angioblasts in wild-type embryos (A, C) and smo mutants (B, D). (E, F) Confocal microscopy analysis revealing the normal size of the dorsal aorta, the posterior cardinal vein and intersomitic vessels in control embryos (E), and an expanded posterior cardinal vein in cyclopamine-treated embryos (F). (G) Schematic representation depicting the temporal activity of cyclopamine in artery–vein specification. Embryos were incubated in 50 μM cyclopamine during a series of developmental window. Red and blue line: artery and vein. Blue line: vein. 5 s:5-somite stage. 10 s:10-somite stage. |
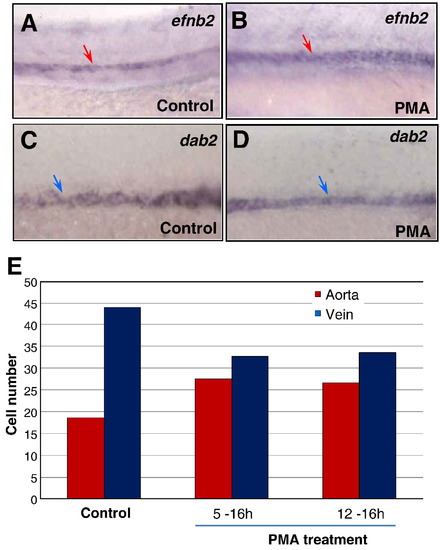
Activated Hh signaling causes an increase of arterial cells and loss of venous cells. (A–D) In situ hybridization analysis displaying the increased efnb2 expression in purmorphamine (PMA)-treated embryos (A) compared to controls (B); and the reduced dab2 expression in embryos treated with PMA (C) compared to controls (0.1% DMSO) (D). (E) Bar chart depicting numbers of arterial and venous cells in PMA-treated embryos from 5 to 16 hpf and 12 to 16 hpf, compared to controls (0.1% DMSO). Arterial cell number: 19.3 ± 0.6 (control); 27.5 ± 0.9 (5–16 hpf; PMA treated); 26.7 ± 0.6 (12–16 hpf; PMA treated). Venous cell number: 41.6 ± 1.5 (control); 32.7 ± 1.2 (5–16 hpf; PMA treated); 33.5 ± 1.8 (12–16 hpf; PMA treated). Endothelial cells were counted from 6 sections derived from 3 wild-type embryos or 3 PMA-treated embryos. Each section covers four segment lengths of intersomitic vessels along the dorsal aorta and the posterior cardinal vein in the middle trunk above the yolk extension region. a: aorta. v: vein. |
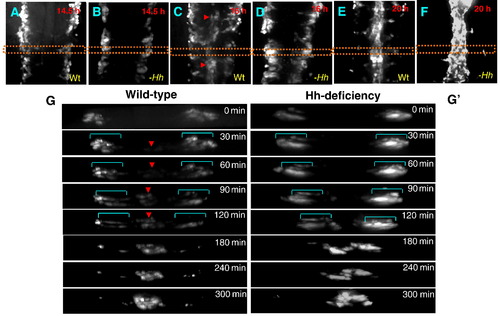
The first wave of angioblast migration is diminished in embryos with Hh signaling deficiency. (A–F) Time-lapse imaging revealing sequential migration of angioblasts toward the midline from 14.5 hpf to 20 hpf in wild-type embryos (A;C;E), and cyclopamine-treated embryos (B;D;F). (G and G′) Crossover optical sections derived from the time-lapse sequence viewing angioblast migration in wild-type embryos (G), and cyclopamine-treated embryos (G′). Red arrowhead: arterial cells. Blue bracket: venous cells. Frames were captured from 300-min time-lapse sequence. Images were obtained from the dorsal side of transgenic embryos Tg(fli1:EGFP) (A–F). The 0 min time point is from the approximately 14.5 hpf stage (G;G′). Times shown in right corners represent time elapsed after the start of the sequence (G;G′). Yellow dot lines (A–F) indicate where the optical crossover sections in G and G′ were derived from. |
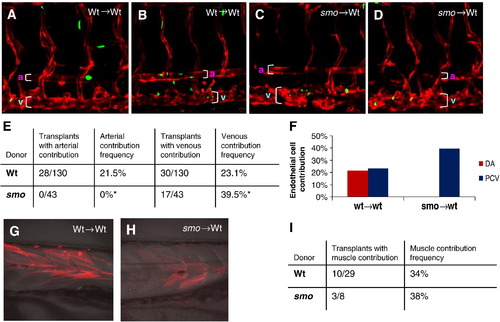
Hh signaling acts cell-autonomously to induce arterial cell formation by inhibiting venous cell development. (A, B) Confocal microscopic analysis revealing the presence of EGFP-positive donor cells derived from wild-type Tg(fli:EGFP-nuc) embryos in the dorsal aorta and the posterior cardinal vein of wild-type host embryos Tg(flk:DsRed) at 72 hpf, when the dorsal aorta is completely separated from the cardinal vein. (C, D) Confocal optics showing the presence of EGFP-positive smo-null cells derived from heterozygous crosses of smo mutants [smo -/+; Tg(fli:EGFP-nuc)] in the posterior cardinal vein in wild-type host embryos Tg(flk:DsRed) at 72 hpf. (E) Data table quantitating transplantation results. A ratio of hosts with donor-derived arterial (or venous) cells to all host embryos with donor cells in any tissue was used to determine arterial (or venous) contribution frequency. (F) Bar chart depicting arterial or venous contribution frequency. (G, H) Fluorescent optics showing transplantation of fluorescein-dextran injected donor cells into wild-type hosts. Both wild-type and smo donors give rise to skeletal muscle tissue. (I) Data table quantitating transplantation results of fluorescein-dextran injected hosts. A ratio of hosts with donor-derived muscle cells to all host embryos was used to determine muscle contribution frequency. |
Reprinted from Developmental Biology, 341(1), Williams, C., Kim, S.H., Ni, T.T., Mitchell, L., Ro, H., Penn, J.S., Baldwin, S.H., Solnica-Krezel, L., and Zhong, T.P., Hedgehog signaling induces arterial endothelial cell formation by repressing venous cell fate, 196-204, Copyright (2010) with permission from Elsevier. Full text @ Dev. Biol.