
- Title
-
Environmental and genetic modifiers of squint penetrance during zebrafish embryogenesis
- Authors
- Pei, W., Williams, P.H., Clark, M.D., Stemple, D.L., and Feldman, B.
- Source
- Full text @ Dev. Biol.
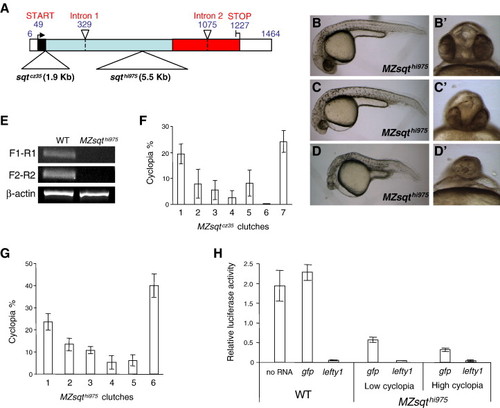
Genetic background influences sqt penetrance. (A) Schematic diagram of sqtcz35 and sqthi975 alleles. The following features of sqt's three exons are indicated: 5′ and 3′ UTR (white boxes); signal sequence (black box); pro-domain (light blue box) and mature ligand domain (red box). Points of the sqtcz35 1.9 Kb and the sqthi975 5.5 Kb insertions (large triangles) and introns (small triangles, dashed lines) are indicated. (B–D) Phenotypic range of MZsqthi975/hi95embryos. Lateral views of normal (B), mildly affected (C) and strongly affected (D) MZsqthi975/hi975 embryos. Apostrophes (′) indicate a frontal view of the same specimen. (E) RT-PCR analysis of sqt mRNA in MZsqthi975/hi975embryos. cDNA extracted from WT and MZsqthi975/hi975 embryos at the 8-cell stage were used as template to analyze maternal sqt mRNA levels. F1-R1 shows the amplicon obtained from F1 and R1 primer pair, spanning the mRNA coordinates 3 to 165. F2-R2 shows the amplicon obtained from F2 and R2 primer pair, spanning the mRNA coordinates 962–1180. β-actin was used as an internal reference for the RT-PCR. Both amplicons are reduced or absent in MZsqthi975/hi975 embryos. (F–G) Genetic background determines sqt penetrance. For both MZsqtcz35/cz35 (F) and MZsqthi975/hi975 (G) embryos, incidences of cyclopia were scored on the second day of development (30–36 hpf). Reproducible cyclopia penetrance rates were observed over repeat crosses of the same parental pairs, whereas distinct penetrance rates were often obtained from different crosses. Each bar represents the average cyclopia penetrance of several hundred (197 < n < 774) embryos from three (MZsqtcz35/cz35) or four (MZsqthi975/hi975) repeat crosses, with standard deviations indicated. See Table S1 for details. (H) Residual Nodal activity is inversely correlated with cyclopia penetrance. WT embryos, as well as low-cyclopia (2%, n = 112) and high-cyclopia (24%, n = 107) MZsqthi975/hi975 embryos were used to measure the Nodal activity by ARE-reporter assay. 25 pg of luciferase reporter plasmids, plus 30 pg of either gap43-gfp or lefty1 mRNA were injected into each group of embryos. WT embryos without mRNA injection (labeled as “no RNA”) were used to establish standard Nodal activity levels. WT embryos injected with lefty1 mRNA were used to establish the baseline activity of the reporter system in the absence of Nodal activity. Average values and standard deviations of triplicate experiments are shown. PHENOTYPE:
|
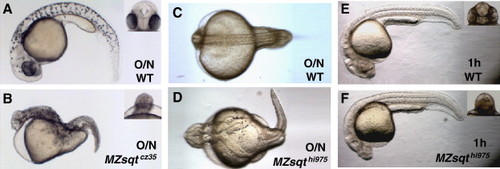
Heat shifts induce dysmorphology in Sqt-deficient embryos. (A–D) Overnight heat treatment produces severe dysmorphology in MZsqt embryos. Incubation at 34 °C overnight caused relatively mild defects in WT embryos (A, C) but dramatically increased phenotypic expressivity in MZsqt-/- embryos (B, D). The loss of anterior mesendoderm derivatives and severe cyclopia in the heated MZsqtcz35/cz35 embryo shown in 2B resembles sqt-/-;cyc-/- or sqt-/-;cyc+/- compound mutant phenotypes ([Dougan et al., 2003] and [Feldman et al., 1998]). Partial bifurcations of the midline, as shown in Fig. 2D, were often seen in MZsqt-/- embryos that were heated overnight. (E–F) 1 h heat treatment during the post-MBT blastula stage exclusively induces cyclopia in MZsqt embryos. 1 h heat treatment of WT or MZsqt-/- embryos (E, F), did not produce any novel abnormalities, but did increase the incidence of cyclopia in MZsqt-/- embryos. |
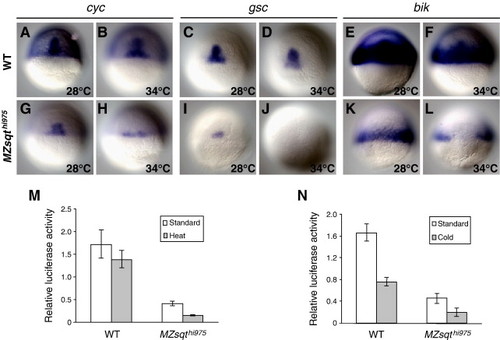
Temperature affects Nodal activity. (A–L) Temperature increase causes reductions in Nodal target gene expression. WT (A–F) and MZsqthi975/hi975 (G–L) embryos from single respective clutches were incubated either at 28 °C from 0 to 6 h (A, C, E, G, I, K) or 28 °C from 0–1.75 h and 34 °C from 1.75 to 5.3 h (B, D, F, H, J, L), then collected and fixed when shield formation was apparent and probed for cyc (A,B,G,H) ([Rebagliati et al., 1998a] and [Sampath et al., 1998]), goosecoid (C,D,I,J) (Stachel et al., 1993) or bhikhari (E,F,K,L) (Vogel and Gerster, 1999) expression by whole-mount in situ hybridization. (M–N) Temperature shifts cause reductions in Nodal activity. WT and MZsqthi975/hi975 embryos from single respective clutches were injected with 25 pg of luciferase reporter plasmids, incubated at 34 °C or 15 °C for 1 h starting at the 256-cell stage (2.5 hpf), or kept at 28 °C, then collected at shield stage and measured for luciferase activity. Significant reductions in activity were seen in MZsqthi975/hi975 embryos subjected to heat or cold, as well as WT embryos subjected to cold. EXPRESSION / LABELING:
|
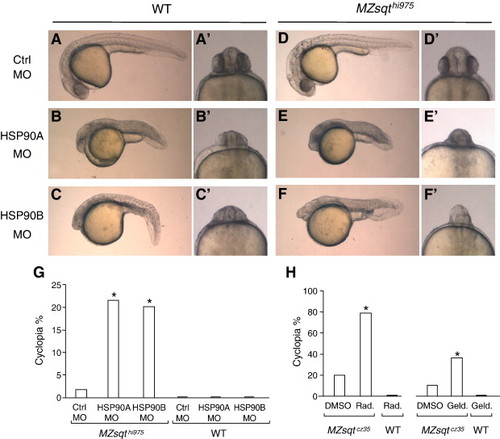
Inhibition of HSP90 function increases cyclopia penetrance in Sqt-deficient embryos. (A–F) Phenotypes from MO knockdown of either HSP90A or HSP90B in WT and MZsqthi975/hi975 embryos. WT and MZsqthi975/hi975 embryos were injected with 2 ng of either control MO, HSP90A MO or HSP90B MO. All injected embryos were incubated overnight and scored for cyclopia on day 2. Apostrophes (′) indicate a frontal view of the same specimen. A variety of defects, including small heads, short axes and compressed, U-shaped somites were observed both in MZsqt and WT embryos (B, C, E, F). In addition, both HSP90A MO and HSP90B MO increased the incidence of cyclopia specifically in MZsqt, but not WT, embryos (E′, F′, G). (G) MO knockdown of either HSP90A or HSP90B increases the cyclopia rate in MZsqthi975/hi975 embryos. The number of WT embryos injected and scored for cyclopia as above were: Ctrl MO, n = 41; HSP90A MO, n = 35; HSP90B MO, n = 39. The MZsqthi975/hi975 embryo numbers were: Ctrl MO, n = 57; HSP90A MO, n = 60; HSP90B MO, n = 74. The observed differences in cyclopia rates had P < 0.001 and P = 0.001 chances of occurring randomly, for the HSP90A and HSP90B experiments, respectively. (H) HSP90-inhibitory drugs induce cyclopia in MZsqtcz35/cz35 embryos. To test the effects of drugs inhibiting HSP90, WT and MZsqtcz35/cz35 embryos were injected with radicicol (WT, n = 19; MZsqt, n = 19), geldanamycin (WT, n = 18; MZsqt, n = 46) or carrier buffer (DMSO) alone (WT, n = 19; MZsqt, n = 49). All injected embryos were incubated overnight and scored for cyclopia on day 2. The observed differences in cyclopia rates had P < 0.001 and P = 0.008 chances of occurring randomly, for the radicicol and geldanamaycin experiments, respectively. PHENOTYPE:
|
Reprinted from Developmental Biology, 308(2), Pei, W., Williams, P.H., Clark, M.D., Stemple, D.L., and Feldman, B., Environmental and genetic modifiers of squint penetrance during zebrafish embryogenesis, 368-378, Copyright (2007) with permission from Elsevier. Full text @ Dev. Biol.